Small Molecule Inhibitors Targeting Gαi2 Protein Attenuate Migration of Cancer Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Gαi2 Inhibitors Design: Molecular Docking Analysis and Synthesis
2.2. Inhibition of Gαi2 Activation Decreases the Migration and Invasion in Prostate Cancer Cells
2.3. Compound 14 Blocks Activation of Gαi2
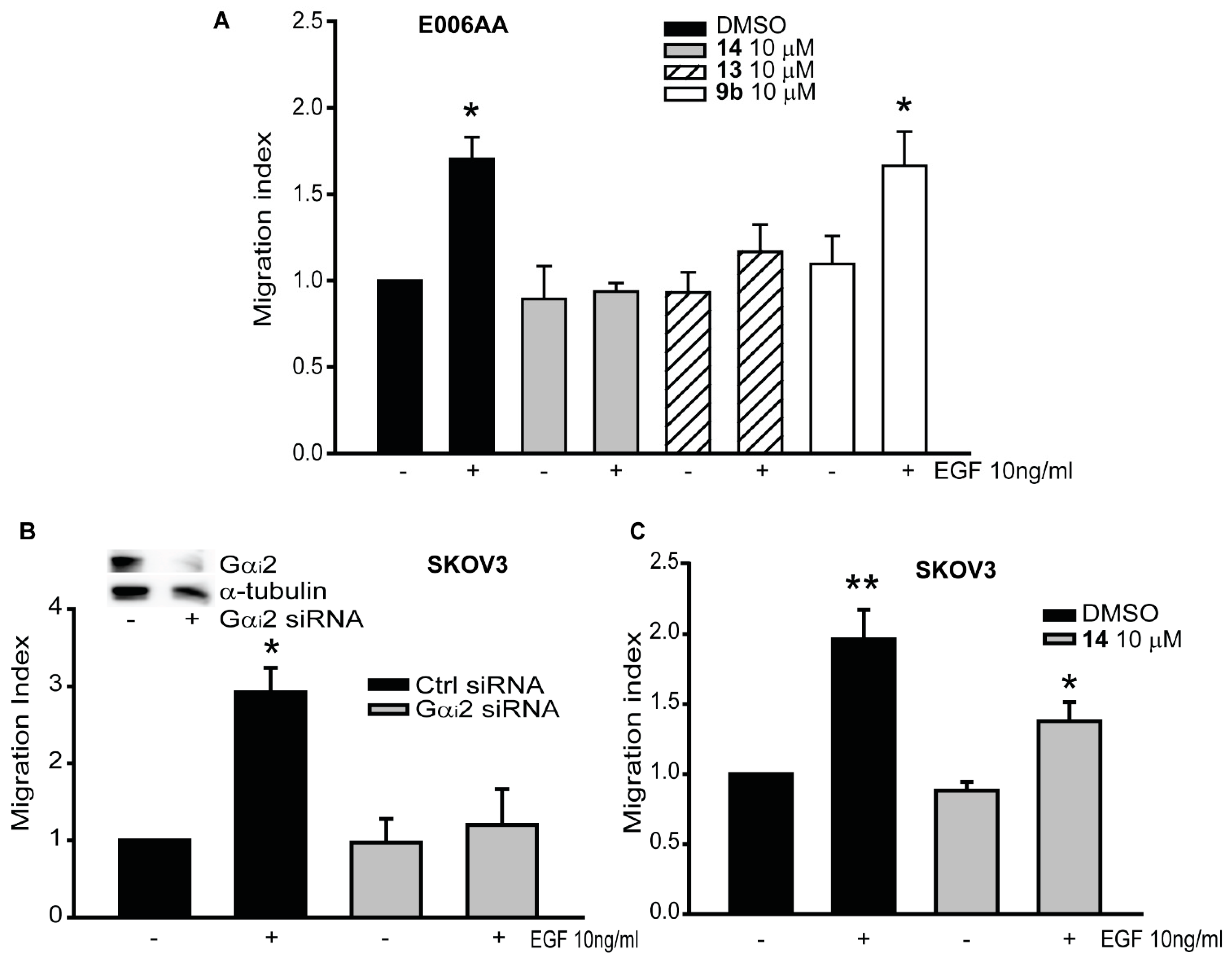
2.4. Gαi2 Protein is Essential for Cell Migration in Renal and Ovarian Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Small Molecules Preparation and Docking
4.4. Immunoprecipitation of Active Gαi
4.5. Transient Transfection with Constitutively Active Gαi2-Q205L Plasmid
4.6. Western Blot Analysis
4.7. Cell Viability Assay
4.8. Cell Migration and Invasion Assays
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Sokeland, G.; Schumacher, U. The functional role of integrins during intra- and extravasation within the metastatic cascade. Mol. Cancer 2019, 18, 12. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massague, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [Green Version]
- Appert-Collin, A.; Hubert, P.; Cremel, G.; Bennasroune, A. Role of ErbB Receptors in Cancer Cell Migration and Invasion. Front. Pharmacol. 2015, 6, 283. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Knaut, H. Chemokine signaling in development and disease. Development 2014, 141, 4199–4205. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Wu, D.; Xu, B.; Qian, W.; Li, P.; Lu, Q.; Yin, C.; Zhang, W. microRNA-133 inhibits cell proliferation, migration and invasion in prostate cancer cells by targeting the epidermal growth factor receptor. Oncol. Rep. 2012, 27, 1967–1975. [Google Scholar] [CrossRef]
- Meirson, T.; Gil-Henn, H. Targeting invadopodia for blocking breast cancer metastasis. Drug Resist. Updates 2018, 39, 1–17. [Google Scholar] [CrossRef]
- Bar-Shavit, R.; Maoz, M.; Kancharla, A.; Nag, J.K.; Agranovich, D.; Grisaru-Granovsky, S.; Uziely, B. G Protein-Coupled Receptors in Cancer. Int. J. Mol. Sci. 2016, 17, 1320. [Google Scholar] [CrossRef] [Green Version]
- Decaillot, F.M.; Kazmi, M.A.; Lin, Y.; Ray-Saha, S.; Sakmar, T.P.; Sachdev, P. CXCR7/CXCR4 heterodimer constitutively recruits beta-arrestin to enhance cell migration. J. Biol. Chem. 2011, 286, 32188–32197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duc, N.M.; Kim, H.R.; Chung, K.Y. Recent Progress in Understanding the Conformational Mechanism of Heterotrimeric G Protein Activation. Biomol. Ther. (Seoul) 2017, 25, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, L.; Fu, H.; Yan, J.; Wang, Y.; Guo, H.; Hao, X.; Xu, X.; Jin, T.; Zhang, N. Association between Galphai2 and ELMO1/Dock180 connects chemokine signalling with Rac activation and metastasis. Nat. Commun. 2013, 4, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, P.; Garcia-Marcos, M.; Bornheimer, S.J.; Farquhar, M.G. Activation of Galphai3 triggers cell migration via regulation of GIV. J. Cell Biol. 2008, 182, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Clarke, S.; Vo, B.T.; Khan, S.A. The essential role of Gialpha2 in prostate cancer cell migration. Mol. Cancer Res. 2012, 10, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Zhong, M.; Boseman, M.L.; Millena, A.C.; Khan, S.A. Oxytocin induces the migration of prostate cancer cells: Involvement of the Gi-coupled signaling pathway. Mol. Cancer Res. 2010, 8, 1164–1172. [Google Scholar] [CrossRef] [Green Version]
- Caggia, S.; Chunduri, H.; Millena, A.C.; Perkins, J.N.; Venugopal, S.V.; Vo, B.T.; Li, C.; Tu, Y.; Khan, S.A. Novel role of Gialpha2 in cell migration: Downstream of PI3-kinase-AKT and Rac1 in prostate cancer cells. J. Cell. Physiol. 2018, 234, 802–815. [Google Scholar] [CrossRef]
- Banon-Rodriguez, I.; Saez de Guinoa, J.; Bernardini, A.; Ragazzini, C.; Fernandez, E.; Carrasco, Y.R.; Jones, G.E.; Wandosell, F.; Anton, I.M. WIP regulates persistence of cell migration and ruffle formation in both mesenchymal and amoeboid modes of motility. PLoS ONE 2013, 8, e70364. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Abel, P.W.; Kirui, J.K.; Deng, C.; Sharma, P.; Wolff, D.W.; Toews, M.L.; Tu, Y. Identification of upregulated phosphoinositide 3-kinase gamma as a target to suppress breast cancer cell migration and invasion. Biochem. Pharmacol. 2013, 85, 1454–1462. [Google Scholar] [CrossRef] [Green Version]
- Burns, D.L. Subunit structure and enzymic activity of pertussis toxin. Microbiol. Sci. 1988, 5, 285–287. [Google Scholar]
- Appleton, K.M.; Bigham, K.J.; Lindsey, C.C.; Hazard, S.; Lirjoni, J.; Parnham, S.; Hennig, M.; Peterson, Y.K. Development of inhibitors of heterotrimeric Gα(i) subunits. Bioorg. Med. Chem. 2014, 22, 3423–3434. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malkov, A.V.; Vrankova, K.; Stoncius, S.; Kocovsky, P. Asymmetric reduction of imines with trichlorosilane, catalyzed by sigamide, an amino acid-derived formamide: Scope and limitations. J. Org. Chem. 2009, 74, 5839–5849. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Sun, L.; Jiao, Y.; Lee, L.T.O. The Role of G Protein-coupled Receptor Kinases in Cancer. Int. J. Biol. Sci. 2018, 14, 189–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, B.; Gschwind, A.; Ullrich, A. Multiple G-protein-coupled receptor signals converge on the epidermal growth factor receptor to promote migration and invasion. Oncogene 2003, 23, 991. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.G.; Sanders, A.J.; Katoh, M.; Ungefroren, H.; Gieseler, F.; Prince, M.; Thompson, S.K.; Zollo, M.; Spano, D.; Dhawan, P.; et al. Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin. Cancer Biol. 2015, 35, S244–S275. [Google Scholar] [CrossRef]
- Arnst, J.L.; Hein, A.L.; Taylor, M.A.; Palermo, N.Y.; Contreras, J.I.; Sonawane, Y.A.; Wahl, A.O.; Ouellette, M.M.; Natarajan, A.; Yan, Y. Discovery and characterization of small molecule Rac1 inhibitors. Oncotarget 2017, 8, 34586–34600. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Zheng, Y. Approaches of targeting Rho GTPases in cancer drug discovery. Expert Opin. Drug Discov. 2015, 10, 991–1010. [Google Scholar] [CrossRef] [Green Version]
- Entschladen, F.; Zanker, K.S.; Powe, D.G. Heterotrimeric G protein signaling in cancer cells with regard to metastasis formation. Cell Cycle 2011, 10, 1086–1091. [Google Scholar] [CrossRef]
- Hoelder, S.; Clarke, P.A.; Workman, P. Discovery of small molecule cancer drugs: Successes, challenges and opportunities. Mol. Oncol. 2012, 6, 155–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venugopal, S.V.; Caggia, S.; Gambrell-Sanders, D.; Khan, S.A. Differential roles and activation of mammalian target of rapamycin complexes 1 and 2 during cell migration in prostate cancer cells. Prostate 2020, 80, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Fu, X.; Chen, P.; Wu, P.; Fan, X.; Li, N.; Zhu, H.; Jia, T.T.; Ji, H.; Wang, Z.; et al. SPSB1-mediated HnRNP A1 ubiquitylation regulates alternative splicing and cell migration in EGF signaling. Cell Res. 2017, 27, 540–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, B.T.; Morton, D., Jr.; Komaragiri, S.; Millena, A.C.; Leath, C.; Khan, S.A. TGF-beta effects on prostate cancer cell migration and invasion are mediated by PGE2 through activation of PI3K/AKT/mTOR pathway. Endocrinology 2013, 154, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Vo, B.T.; Khan, S.A. Expression of nodal and nodal receptors in prostate stem cells and prostate cancer cells: Autocrine effects on cell proliferation and migration. Prostate 2011, 71, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Elliott, B.; Zackery, D.L.; Eaton, V.A.; Jones, R.T.; Abebe, F.; Ragin, C.C.; Khan, S.A. Ethnic Differences in TGFbeta Signaling Pathway May Contribute to Prostate Cancer Health Disparity. Carcinogenesis 2018. [Google Scholar] [CrossRef]
- Koochekpour, S.; Maresh, G.A.; Katner, A.; Parker-Johnson, K.; Lee, T.J.; Hebert, F.E.; Kao, Y.S.; Skinner, J.; Rayford, W. Correction: Establishment and characterization of a primary androgen-responsive African-American prostate cancer cell line, E006AA. Prostate 2019, 79, 815. [Google Scholar] [CrossRef] [Green Version]
- Sammond, D.W.; Eletr, Z.M.; Purbeck, C.; Kimple, R.J.; Siderovski, D.P.; Kuhlman, B. Structure-based protocol for identifying mutations that enhance protein-protein binding affinities. J. Mol. Biol. 2007, 371, 1392–1404. [Google Scholar] [CrossRef] [Green Version]
- Gryder, B.E.; Rood, M.K.; Johnson, K.A.; Patil, V.; Raftery, E.D.; Yao, L.P.; Rice, M.; Azizi, B.; Doyle, D.F.; Oyelere, A.K. Histone deacetylase inhibitors equipped with estrogen receptor modulation activity. J. Med. Chem. 2013, 56, 5782–5796. [Google Scholar] [CrossRef] [Green Version]
- Raji, I.; Yadudu, F.; Janeira, E.; Fathi, S.; Szymczak, L.; Kornacki, J.R.; Komatsu, K.; Li, J.D.; Mrksich, M.; Oyelere, A.K. Bifunctional conjugates with potent inhibitory activity towards cyclooxygenase and histone deacetylase. Bioorg. Med. Chem. 2017, 25, 1202–1218. [Google Scholar] [CrossRef] [Green Version]
- Vo, B.T.; Cody, B.; Cao, Y.; Khan, S.A. Differential role of Sloan-Kettering Institute (Ski) protein in Nodal and transforming growth factor-beta (TGF-beta)-induced Smad signaling in prostate cancer cells. Carcinogenesis 2012, 33, 2054–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caggia, S.; Tapadar, S.; Wu, B.; Venugopal, S.V.; Garrett, A.S.; Kumar, A.; Stiffend, J.S.; Davis, J.S.; Oyelere, A.K.; Khan, S.A. Small Molecule Inhibitors Targeting Gαi2 Protein Attenuate Migration of Cancer Cells. Cancers 2020, 12, 1631. https://doi.org/10.3390/cancers12061631
Caggia S, Tapadar S, Wu B, Venugopal SV, Garrett AS, Kumar A, Stiffend JS, Davis JS, Oyelere AK, Khan SA. Small Molecule Inhibitors Targeting Gαi2 Protein Attenuate Migration of Cancer Cells. Cancers. 2020; 12(6):1631. https://doi.org/10.3390/cancers12061631
Chicago/Turabian StyleCaggia, Silvia, Subhasish Tapadar, Bocheng Wu, Smrruthi V. Venugopal, Autumn S. Garrett, Aditi Kumar, Janae S. Stiffend, John S. Davis, Adegboyega K. Oyelere, and Shafiq A. Khan. 2020. "Small Molecule Inhibitors Targeting Gαi2 Protein Attenuate Migration of Cancer Cells" Cancers 12, no. 6: 1631. https://doi.org/10.3390/cancers12061631